
Characteristics of Soil Active Carbon Fractions and Carbon Pool Stability in Major Forest Types in Northern Daxing'an Mountains
-
摘要:
目的 土壤有机碳是森林生态系统碳循环的重要组成部分, 活性有机碳能够指示土壤碳库变化, 研究不同森林类型土壤活性碳组分及碳库稳定性的变化规律对理解碳循环的生物地球化学过程有重要意义, 可为面向固碳减排的生态系统碳库管理提供依据。 方法 以大兴安岭北部兴安落叶松林、樟子松林、白桦林和山杨林为研究对象, 于2023年5—9月采集0—10, 10—20 cm土层土壤样品, 采用改良后的Loginow法测定土壤中活性易氧化碳含量, 利用活性碳利用率(ER)及碳库活度(CPA)分析不同林型下土壤碳库稳定性差异。 结果 (1) 5—9月, 4种林型土壤总有机碳(TOC)、高活性易氧化碳(LOC-H)和中活性易氧化碳(LOC-M)均呈"单峰"变化趋势, 于8月达到峰值, 且阔叶林含量高于针叶林。(2)土壤稳定性有机碳利用率(ER4)在5—9月为72.33%~85.11%, 显著高于其他3种活性碳利用率, 在土壤碳库中占主导地位。(3)土壤碳库活度(CPA)为0.20~0.38, 且山杨林>白桦林>樟子松林>兴安落叶松林, 针叶林于8月达到峰值, 阔叶林则在5月最高。(4)土壤活性有机碳含量和碳库稳定性受气候因素和土壤因素的共同作用。蔗糖酶为土壤因子中主要影响因素, 而气候因子中降雨量和光合有效辐射影响较大。寒温带针叶林土壤有机碳整体稳定性大于阔叶林, 但在温度较高情况下, 针叶林深层土壤损失有机碳速率高于阔叶林。 结论 研究结果有助于提高对寒温带森林土壤活性碳及有机碳库的认识, 建议该地区尽快人工促进次生林演替或引入针叶树种, 使其形成混交林, 但在全球日益变暖的背景下, 阔叶林在土壤碳库中发挥的作用也不容忽视。 Abstract:Objective Soil organic carbon is an important component of the carbon cycle in forest ecosystems, and active organic carbon can indicate changes in soil carbon pools. Studying the pattern of change of soil active carbon fractions and carbon pool stability in different forest types is of great significance to the understanding of the biogeochemical processes of the carbon cycle, and can provide a basis for the management of ecosystem carbon pools oriented to carbon sequestration and emission reduction. Methods The soil samples of 0-10 and 10-20 cm soil layers were collected from May to September in the growing season from Larix gmelinii, Pinus sylvestris var. mongolica, Betula platyphylla and Populus davidiana forests in the northern part of Daxing' anling, and the contents of different reactive carbon oxides in the soils were measured by the modified Loginow method, and the active carbon utilisation (ER) and carbon pool activity (CPA) were used to measure the carbon pool stability and the carbon cycle. Results (1) From May to September, soil total organic carbon (TOC), highly active organic carbon (LOC-H) and moderately active organic carbon (LOC-M) of the four forest types showed a "single-peak" trend of change, and reached a peak in August, and the contents were higher in broad-leaved forests than in coniferous forests. (2) The utilisation rate of soil stability organic carbon (ER4) fluctuated between 72.33% and 85.11% from May to September, it was significantly higher than the utilisation rates of the other three types of active carbon, and dominated the soil carbon pool. (3) Soil carbon pool activity (CPA) fluctuated between 0.20 and 0.38, with P. davidiana forests>B. platyphylla forests>P. sylvestris var. mongolica forests >L. gmelinii forests; that of coniferous forests peaked in August, while that of broad-leaved forests were highest in May. (4) Soil active organic carbon content and carbon pool stability were subject to the combined effects of climatic factors and soil factors. Sucrase was the main influence of soil factors, while rainfall and photosynthetically active radiation were more influential among climatic factors. The overall stability of soil organic carbon in coniferous forests in the cold temperate zone was greater than that in broad-leaved forests, but the rate of organic carbon loss from deeper soil layers in coniferous forests was higher than that in broad-leaved forests under higher temperatures. Conclusion This study helps to improve the understanding of soil active carbon and organic carbon pools in cold temperate forests, artificially promoting secondary forest succession or introducing coniferous species as soon as possible is suggested to form mixed forests in the region, but the role played by broad-leaved forests in soil carbon pools should not be ignored in the context of increasing global warming. -
“碳达峰,碳中和”已成为本世纪中叶我国生态环境建设的重要战略目标[1]。森林因其较高的物质生产能力和巨大的碳储存能力,在“碳中和”目标实现过程中被寄予了厚望。土壤有机碳库是大气重要的“源”和“汇”,在全球碳循环中起着至关重要的作用,是实现“双碳”目标的关键一环[2]。由于土壤碳库基数巨大,TOC难以灵敏反映短期内整个碳库的微小变化,因此有研究[3-4]将TOC分为活性有机碳(LOC)和稳定性有机碳(NLOC),并指出土壤有机碳的短期变化主要发生在LOC中。以往对于LOC组分的测定大多是从物理、化学和生物层面进行考虑,但由于其测定方法和提取溶剂存在较大差异,使三者并非完全独立,部分活性碳组分间存在重合,而改良过的LOGNINOW法[5]通过使用不同浓度的KMnO4氧化后进行差减,将LOC分为高活性、中活性和低活性3种组分,既规避了不同溶剂引起的差别,同时又保证各组分间相互独立[6],其不同组分在质、量上的变化,能够相对全面反映土壤有机碳库积累程度及其稳定性[7]。
土壤有机碳稳定性是评价土壤长期固碳潜力的重要指标,是由多种环境因素共同作用所决定的[8]。DEVI[9]通过对国内外55种林型土壤有机碳含量及影响因素进行汇总发现,针叶林土壤固碳能力高于阔叶林,其中年平均温(MAT)和砂粉含量为主导因素;SU等[10]对鼎湖山生物保护区常绿针叶林和阔叶林土壤活性碳研究发现,阔叶林向针叶林转变后通过影响土壤化学性质、微生物生物量及大团聚体的形成,使土壤碳固存能力低于针叶林;贾匡迪等[11]对山西太岳山典型针叶林土壤活性碳组分进行研究发现,油松林土壤有机碳稳定性显著低于华北落叶松林,土壤含水率和土壤酶活性是主要影响因子。由此可知,植被类型、气候类别和土壤属性在不同区域上共同影响着土壤活性有机碳及碳库稳定性。
有学者[12]研究指出,土壤有机碳含量随纬度的增加呈指数增长模式,北半球高纬度地区土壤碳库研究逐渐成为生物圈CO2通量研究的潜在热点。大兴安岭北部作为我国唯一的高纬度寒温带林区,气候严寒,生长季短,昼夜温差大,是多年冻土的主要分布区,同时也是对全球气温变化反应最敏感的地区之一[13]。因此,对寒温带森林土壤活性碳组分及碳库稳定性的深入研究是十分必要的。前人对大兴安岭林区土壤碳库进行了大量研究,但多集中于碳储量[14]和团聚体稳定性[15]方面,且多关注于兴安落叶松林[8],关于该林区4种典型森林土壤活性碳组分及碳库稳定性的研究相对匮乏。在全球温室效应日益严重和大气CO2浓度不断升高的背景下,深入解析该林区不同林型土壤活性碳组分含量及影响因素,可为进一步挖掘森林土壤碳汇潜力,更高效地发挥土壤碳库作用提供科学理论依据,对如期实现我国“碳达峰,碳中和”目标具有重要意义。
基于此,本研究以大兴安岭寒温带4种典型森林类型(兴安落叶松林、樟子松林、白桦林和山杨林)为研究对象,通过探讨其土壤活性碳组分含量动态变化规律,探究不同林型下土壤有机碳的稳定性,明确影响该地区土壤活性碳及碳库稳定的气候因子和土壤因素,以期为寒温带森林生态系统碳循环及碳稳定性研究提供科学参考。
1. 研究区概况与研究方法
1.1 研究区概况
研究区选择在黑龙江漠河森林生态系统国家定位观测研究站,位于大兴安岭北部漠河县北极村内(123°15′30″E,53°33′30″N)。该地区属寒温带大陆性季风气候,年平均气温-4.9 ℃,年降水量约400 mm,无霜期80~90天,降水多集中在夏季。全年冰雪覆盖时间最长可达200天。地带性土壤以棕色针叶林土为主,局部地区还有草甸土和沼泽土分布,是我国多年冻土的主要分布区。森林植被系欧亚大陆寒温带明亮针叶林,以兴安落叶松(Larix gmelinii)林为优势建群种,此外还有樟子松(Pinus sylvestris var. mongolica)林、白桦(Betula platyphylla)林和山杨(Populus davidiana) 林等森林类型,林下灌木主要有兴安杜鹃(Rhododendron dauricum)、杜香(Rhododendron tomentosum)、笃斯越橘(Vaccinium uliginosum)等。观测期间相关气候因素见表 1。
表 1 研究区2023年5—9月相关气候因素Table 1 Changes in relevant climatic factors in the study area, from May to September 2023月份 空气温度/℃ 相对湿度/% 降雨量/mm 太阳有效辐射/(W·m-2) 5 9.26 54.87 51.40 107.95 6 15.66 73.52 59.10 131.11 7 20.17 75.86 37.70 135.37 8 16.89 70.71 33.70 112.63 9 8.98 70.07 26.20 56.42 注:表中空气湿度、相对湿度和太阳有效辐射均为当月平均值;降雨量为当月总降雨量。 1.2 样地设置与样品采集
于2023年5月在前期踏查基础上,选择大兴安岭北部4种典型森林类型为研究对象,即兴安落叶松林、樟子松林、白桦林和山杨林。并在各林型典型地段分别设置3块样地,面积为20 m×30 m。对样地进行每木检尺,记录基本信息(表 2)。由于大兴安岭寒温带土层较薄,20 cm以下多含有大量的石块,故本研究仅对0—20 cm土层进行研究。于5—9月在每个样地内以“S”形设置5个采样点,采集土层深度为0—10,10—20 cm土层,土样重约1 kg,去除土壤中的石块和动植物残体,并过2 mm筛备用。将过2 mm筛的新鲜土样分成2部分:一部分土壤用塑封袋密封于冰箱-4 ℃储存,用于测定微生物量碳氮、可溶性碳氮、铵态氮及硝态氮等指标;另一部分土壤于室内风干后测定土壤pH,再取部分土样研磨后过0.149 mm筛后用于测定土壤全碳、活性碳组分及全氮等指标。
表 2 样地基本情况Table 2 Basic information of the sample sites森林类型 海拔/m 林分密度/(株·hm-2) 坡度/(°) 郁闭度 平均胸径/cm 平均树高/m 树种组成 下木种类 LG 305 1 260 6 0.7 14.10±6.2 19.20±5.6 8落1樟1白 1,2,3,4 PS 290 736 10 0.6 21.36±4.9 22.37±3.8 8樟2落 1,3,5 BP 378 1 663 6 0.8 11.50±5.5 13.10±4.1 10白 1,3,4,6 PD 385 975 7 0.7 16.20±6.3 16.50±6.8 8杨2白 6,7,8 注:LG为兴安落叶松林;PS为樟子松林;BP为白桦林;PD为山杨林;DBH为胸径;表中数据为平均值±标准差;1为兴安杜鹃(Rhododen drondauricum);2为杜香(Ledum palustre);3为越桔(Vaccinium vitisidaea);4为笃斯越桔(Vaccinium uliginosum);5为羽节蕨(Gymnocarpium jessoense);6为红花鹿蹄草(Pyrola incarnata);7为舞鹤草(Maianthemum bifolium);8为东方草莓(Fragaria orientalis)。下同。 1.3 测定方法
1.3.1 土壤基本理化性质的测定
土壤含水率(SWC)采用烘干法测定;土壤温度(ST)使用数字式瞬时温度计测定;pH使用PHS-3E型pH计(水土比为2.5∶1)测定;总有机碳(TOC)采用Vario—TOC燃烧法进行测定,全氮(TOC)、铵态氮(NH4+-N)和硝态氮(和NO3--N)使用AA3连续流动分析仪测定,微生物量氮(MBN)采用氯仿熏蒸法进行测定,可溶性有机氮(DON)采用硫酸钾浸提法测定,蔗糖酶(SUC)采用3-5二硝基水杨酸比色法[16]进行测定。
1.3.2 土壤易氧化碳组分的测定
易氧化碳组分(LOC)采用LOGNINOW等[5]提出的KMnO4氧化法提取,并略作改进[6],具体方法为:称取约1.5 g风干土样于50 mL离心管中,加入33.3,167,333 mmol/L KMnO4溶液25 mL,250 r/min振荡1 h,离心5 min(转速4 000 r/min),取上清液按1∶250稀释后于紫外分光光度计565 nm处比色。通过与空白组的吸光值差值计算3种活性碳含量,记为C1、C2、C3(g/Kg,氧化过程中1 mmol/L的高锰酸钾消耗9 mg的碳)。C1为高活性易氧化有机碳(high-labile organic carbon,LOC-H)含量,C2和C1的差值为中活性易氧化有机碳(moderate-labile organic carbon,LOC-M)含量,C3和C2的差值为低活性易氧化有机碳(low labile organic carbon,LOC-L)含量。活性有机碳利用率(ER)、土壤碳库活度(CPA)计算公式为:
$$ \mathrm{NLOC}=\mathrm{TOC}-\mathrm{LOC} $$ (1) $$ E R_1=\frac{\mathrm{LOC}-H}{\mathrm{TOC}} $$ (2) $$ E R_2=\frac{\mathrm{LOC}-M}{\mathrm{TOC}} $$ (3) $$ E R_3=\frac{\mathrm{LOC}-L}{\mathrm{TOC}} $$ (4) $$ E R_4=\frac{\mathrm{TOC}-\mathrm{NLOC}}{\mathrm{TOC}} $$ (5) $$ \mathrm{CPA}=\frac{\mathrm{LOC}}{\mathrm{NLOC}} $$ (6) 式中:LOC为总活性易氧化有机碳含量(g/kg),即C3值;NLOC为稳定性有机碳含量(g/kg);TOC为土壤总有机碳含量(g/kg);CPA为土壤碳库活度;ER1为高活性碳利用率(%);ER2为中活性碳利用率(%);ER3为低活性碳利用率(%);ER4为稳定性碳利用率(%)。
1.4 分析方法
采用Excel 2021对数据进行处理,用SPSS 26.0软件对数据进行统计分析。采用单因素方差法(One way ANOVA)和最小显著性差异法(LSD)检验4种森林类型下土壤活性碳组分的差异显著性(α=0.05);用Canoco 5.0软件对土壤活性碳组分和相关环境因子进行冗余分析(RDA),探究影响该地区土壤碳库稳定的关键环境因子。
2. 结果与分析
2.1 不同森林类型土壤基本理化性质及酶活性动态变化特征
由图 1可知,观测期间研究区4种林型土壤ST、SWC、pH、TN、DON、MBN、NH4+-N、NO3--N和SUC变化趋势略有不同。SWC和DON于5月最高,ST和SUC于8月达到峰值,各土层ST均为樟子松林最高;pH于不同月份间变化幅度较小,且白桦林显著低于其他3种林型(p < 0.05);TN整体呈“双峰”变化,峰值出现于6,8月;MBN和NH4+-N变化趋势相似,均于7,8月相对较高;NO3--N整体呈先升后降再升的波动式变化趋势,且9月显著高于其他月份(p < 0.05)。
 图 1 不同森林类型土壤基本理化性质及酶活性变化特征注:Y1表示左轴柱状图对应的数值,Y2表示右轴折线图对应的数值;图柱上方不同大写字母表示同一林型不同月份间差异达显著水平(p < 0.05);不同小写字母表示同一月份不同林型间差异达到显著水平(p < 0.05)。下同。Fig. 1 Characteristics of changes in basic physical and chemical properties and enzyme activities of soils of different forest types
图 1 不同森林类型土壤基本理化性质及酶活性变化特征注:Y1表示左轴柱状图对应的数值,Y2表示右轴折线图对应的数值;图柱上方不同大写字母表示同一林型不同月份间差异达显著水平(p < 0.05);不同小写字母表示同一月份不同林型间差异达到显著水平(p < 0.05)。下同。Fig. 1 Characteristics of changes in basic physical and chemical properties and enzyme activities of soils of different forest types 下载:
全尺寸图片
下载:
全尺寸图片
2.2 不同森林类型土壤总有机碳(TOC)含量动态变化特征
由图 2可知,研究区5—9月4种林型0—20 cm土层土壤TOC含量为12.80~55.17 g/kg,呈波动式上升至8月最高值后开始下降。0—10 cm土层,樟子松林于5—6月降幅较大,达21.25%,其他3种林型降幅较小,为1.90%~3.76%;6—8月均显著增加,樟子松林增幅最大,达86.43%,兴安落叶松林最小,为21.90%;9月均不同程度降低,但均达显著水平(p < 0.05)。10—20 cm土层,兴安落叶松林和樟子松林于5—6月土壤TOC含量降幅较小,白桦林和山杨林降幅显著(p < 0.05),为33.77%和34.84%;6—8月均显著增加,白桦林增幅达94.78%,樟子松林最小,为13.14%;9月降幅为9.28%~44.60%,但均达显著水平(p < 0.05)。
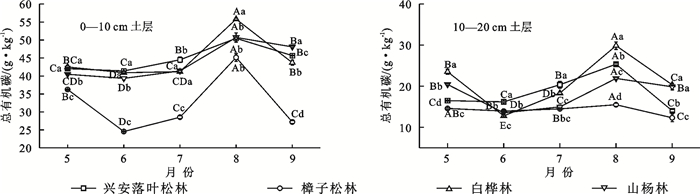 图 2 不同森林类型土壤TOC含量变化特征Fig. 2 Changes of soil total organic carbon contents under different forest types下载:
全尺寸图片
图 2 不同森林类型土壤TOC含量变化特征Fig. 2 Changes of soil total organic carbon contents under different forest types下载:
全尺寸图片
2.3 不同森林类型土壤易氧化碳组分(LOC)含量变化特征
由图 3可知,5—9月,4种林型0—20 cm土层土壤LOC-H、LOC-M和LOC-L含量分别为0.53~4.80,0.59~5.76,0.50~4.79 g/kg,各土层LOC-H、LOC-M均呈先下降后显著上升至8月达到峰值(p < 0.05)。0—10 cm土层为樟子松林增幅最大,分别达289.04%和246.39%,10—20 cm土层为白桦林增幅最大;9月均显著降低(p<0.05),0—10,10—20 cm土层降幅分别为9.28%~44.60%,14.76%~40.17%和40.36%~64.66%,3.17%~65.82%;而LOC-L含量于不同林型、土层和月份间变化趋势各不相同,兴安落叶松林和樟子松林呈波动式上升于7月达到峰值,且此时兴安落叶松林各土层LOC-L含量最高(4.79,1.79 g/kg);而白桦林和山杨林各土层LOC-L含量则于5月最大,6月显著降低后于7月开始上升(p<0.05),0—10 cm土层于8月达到第2个峰值(3.75,4.50 g/kg),而10—20 cm土层则增至7月后逐渐降低(1.49,1.82 g/kg);各林型各土层LOC-L含量均于9月达到最小值,且除兴安落叶松林外,均达显著水平(p<0.05)。
 图 3 不同森林类型土壤易氧化碳组分含量变化特征Fig. 3 Characteristics of changes in the contents of carbon oxidisable fractions in soils of different forest types下载:
全尺寸图片
图 3 不同森林类型土壤易氧化碳组分含量变化特征Fig. 3 Characteristics of changes in the contents of carbon oxidisable fractions in soils of different forest types下载:
全尺寸图片
2.4 不同森林类型土壤活性有机碳利用率(ER)和土壤碳库活度(CPA)变化特征
由图 4可知,观测期间4种林型0—20 cm土层土壤均为ER4显著高于其他3种活性碳利用率(p<0.05),为72.33%~85.11%,表明该地区土壤性质较为稳定。综合5—9月平均值,山杨林0—20 cm土层ER1最高,为3.80%~12.45%,ER2于白桦林最高,为5.08%~10.55%;樟子松林ER1、ER2最低,分别为4.41%~7.55%和4.10%~8.83%,而ER3却大于其他3种林型,为4.28%~13.17%。土壤碳库活度为0.20~0.38(表 3),针叶林于8月达到峰值,阔叶林则在5月最高,0—20 cm土层平均值为山杨林>白桦林>樟子松林>兴安落叶松林。
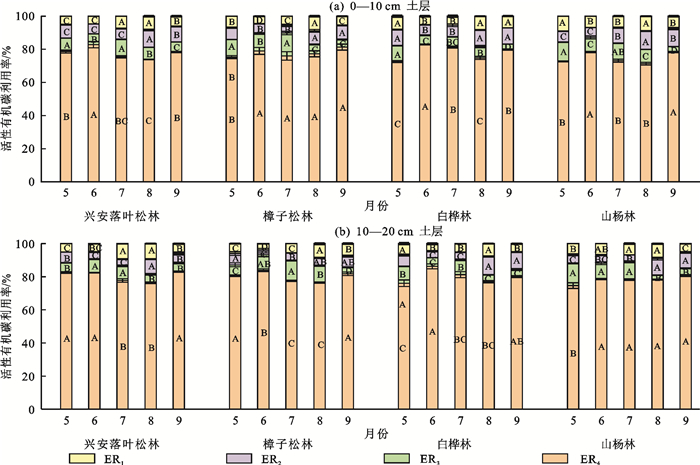 图 4 不同森林类型下土壤活性碳利用率变化特征Fig. 4 Changes of soil active carbon utilization rates under different forest types下载:
全尺寸图片
表 3 不同森林类型下土壤碳库活度变化特征Table 3 Characteristics of changes in soil carbon pool activity under different forest types
图 4 不同森林类型下土壤活性碳利用率变化特征Fig. 4 Changes of soil active carbon utilization rates under different forest types下载:
全尺寸图片
表 3 不同森林类型下土壤碳库活度变化特征Table 3 Characteristics of changes in soil carbon pool activity under different forest types森林类型 土层深度/cm 土壤碳库活度 5月 6月 7月 8月 9月 均值 LG 0-10 0.22±0.01Dc 0.21±0Dc 0.30±0.02Bb 0.35±0.01Ab 0.26±0.03Cb 0.27±0.05 10-20 0.22±0.01Bb 0.21±0Bab 0.29±0.03Aa 0.30±0.01Aa 0.20±0.01Bb 0.25±0.05 PS 0-10 0.30±0.01Ab 0.26±0.03Bb 0.31±0.02Aab 0.32±0.02Ac 0.22±0.01Bc 0.29±0.05 10-20 0.24±0.01Bb 0.22±0.01Cab 0.29±0.01Aa 0.29±0.01Aa 0.21±0.03Cb 0.25±0.04 BP 0-10 0.38±0.01Aa 0.22±0.01Cc 0.26±0.02Bc 0.37±0.01Aa 0.26±0.01Bb 0.30±0.07 10-20 0.32±0.06Aa 0.20±0.02Cb 0.24±0.03BCa 0.28±0.02ABa 0.24±0.02BCab 0.26±0.06 PD 0-10 0.38±0.01Aa 0.30±0.01Ba 0.36±0.01Ba 0.38±0.01Aa 0.31±0.01Ba 0.35±0.04 10-20 0.36±0.01Aa 0.23±0.01Ba 0.24±0.01Ba 0.25±0.01Bb 0.24±0.01Ba 0.27±0.05 注:表中数据为平均值±标准差;同列不同大写字母表示同一林型不同月份间差异达显著水平(p < 0.05);不同小写字母表示同一月份不同林型间差异达到显著水平(p < 0.05)。下同。 2.5 土壤活性碳影响因子分析
以大兴安岭北部4种森林类型0—20 cm土层TOC、LOC-H、LOC-M、LOC-L、ER及CPA作为响应变量,气候因素和土壤基本理化性质作为解释变量进行RDA分析。
由图 5可知,兴安落叶松林第1排序轴和第2排序轴累计解释土壤活性碳特征77.50%的变异,主要影响因子为SUC、QR、MT、TS、TN、ESR和NO3--N,且SUC和QR为主导因子(p=0.002),贡献度率分别为50.4%和26.5%。樟子松林2轴共解释土壤活性碳77.26%的变异,与SUC、ESR、DON、NH4+-N、MBN、和SWC呈显著正相关(p < 0.05),与QR、MT和pH呈显著负相关(p < 0.05),且SUC和ESR贡献度最高,分别达39.00%和18.70%。白桦林和山杨林2轴共解释土壤活性碳79.42%和83.75%变异,土壤主导因子均为SUC和DON(p=0.002),贡献率分别为58.2%,12.6%和54.4%,19.2%,气候主导因子主要为QR和RH(p < 0.05),贡献率分别为5.0%,7.4%和2.4%,1.7%,且除QR、MT和pH外,其他影响因子与3种活性碳含量均呈正相关。
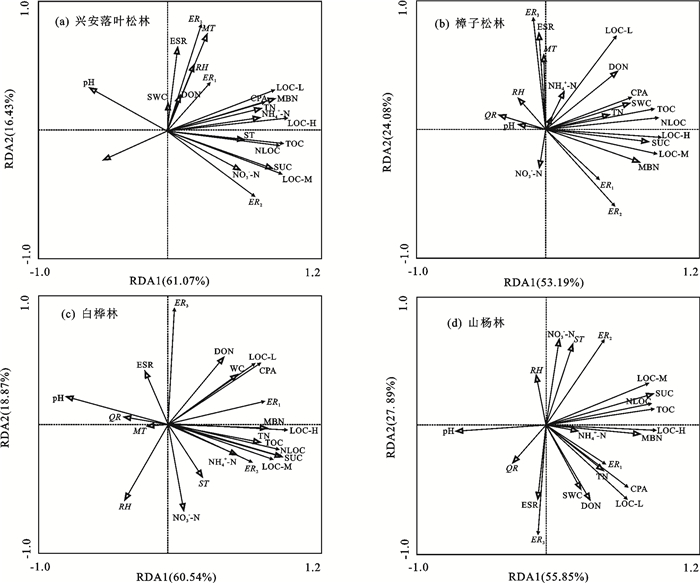 图 5 不同森林类型下土壤活性碳与影响因子冗余分析注:TOC为土壤总有机碳;LOC-H为高活性易氧化有机碳;LOC-M为中活性易氧化有机碳;LOC-L为低活性易氧化有机碳;ER为活性有机碳利用率;CPA为土壤碳库活度;MT为月均温;RH为空气相对湿度;QR为每月总降雨量;ESR为太阳有效辐射;TN为全氮;SUC为蔗糖酶;MBN为微生物氮;DON为可溶性有机氮;NO3--N为硝态氮;NH4+-N为铵态氮;ST为土壤温度;SWC为土壤含水率;pH为土壤酸碱度。Fig. 5 Redundancy analysis of soil active carbon and influencing factors under different forest types下载:
全尺寸图片
图 5 不同森林类型下土壤活性碳与影响因子冗余分析注:TOC为土壤总有机碳;LOC-H为高活性易氧化有机碳;LOC-M为中活性易氧化有机碳;LOC-L为低活性易氧化有机碳;ER为活性有机碳利用率;CPA为土壤碳库活度;MT为月均温;RH为空气相对湿度;QR为每月总降雨量;ESR为太阳有效辐射;TN为全氮;SUC为蔗糖酶;MBN为微生物氮;DON为可溶性有机氮;NO3--N为硝态氮;NH4+-N为铵态氮;ST为土壤温度;SWC为土壤含水率;pH为土壤酸碱度。Fig. 5 Redundancy analysis of soil active carbon and influencing factors under different forest types下载:
全尺寸图片
3. 讨论
3.1 不同森林类型土壤活性碳组分动态变化特征
LOC作为土壤中极不稳定的活性有机碳,分为LOC-H、LOC-M和LOC-L,其中LOC-H包括微生物碳、水溶性碳、可矿化碳等,是土壤中较为活跃的物质,极易受到微生物和环境因子的影响而被植物分解利用[17];而LOC-M、LOC-L包括颗粒有机碳、糖、脂类等物质及部分复杂化合物,如木质素和腐殖质,受外界环境影响相对较小[7],其碳周转速率低于LOC-H。本研究中4种林型各土层LOC-H的变幅大于LOC-M和LOC-L,表明LOC-H对于土壤碳库变化的响应更迅速,与前人[7, 18]研究结果一致。本研究发现,研究区4种林型土壤LOC各组分含量均具有“表聚”特征,是由于植被、枯枝落叶等物质多集中在地表,经微生物分解后形成腐殖质在表层聚集,且植物根系及其分泌物也随土层深度的增加而逐渐减小[19],导致表层含量高于深层。
本研究发现,4种林型各土层LOC-H和LOC-M均表现为8月含量最高,是由于该研究区5月土壤处于解冻期,冰晶体的形成破坏微生物的细胞结构[20],限制土壤中微生物的活动及酶活性;而较低的大气温度(MT)和充足的降雨量(QR)使土壤处于“低温高湿”的环境中,一方面抑制土壤有机碳的矿化速率[21];另一方面为真菌提供适宜的生境,促进各活性碳的积累[22]。5—6月,大气温度回升,冻土融化,土壤中微生物活性提高导致有机碳矿化速率加快[21];同时,冻融交替影响土壤团聚体的稳定性,削弱土壤有机碳的物理保护,DOC的释放进一步促进微生物的活性,土壤活性碳的积累受限[23]。6—8月,植物和微生物进入生长旺季,太阳有效辐射(ESR)的增加促使植物光合作用和代谢速率加快,同时高质量有机物的来源及适宜的水热条件促进微生物的代谢,加快凋落物的分解及根系分泌物的周转速率[24],即使植物、微生物生长过程消耗量较大,但总量仍表现为增加。辜翔等[25]对中亚热带4种林型土壤活性碳季节动态特征研究显示,LOC表现为夏季最高,与本研究结果一致。9月,太阳有效辐射(ESR)降低,植物进入生长末期,根系分泌物随之减少,植物启动自我保护机制,叶片内的养分进行回流保存,凋落物质量降低,微生物的活动受限,各活性碳含量降低[8, 24]。以上分析表明,气候因素通过影响土壤水热条件和植物光合作用间接影响土壤活性有机碳含量,土壤活性碳的波动变化是由气候因子和土壤因素共同作用的结果。
3.2 不同森林类型土壤有机碳稳定性变化特征
ER和CPA在一定程度上可以反映土壤有机碳库的质量,其值越大,表明土壤碳库周转速率越快,土壤碳库越不稳定[19]。本研究发现,研究区4种林型各土层稳定性有机碳利用率为72.33%~85.11%,显著高于史进纳等[26]在亚热带地区的研究结果,却低于高雨[27]在暖温带地区的结论,考虑与本研究区“低温干旱”环境有关。土壤有机碳稳定性与研究区的温度密切相关,温度过高或过低均影响土壤固碳速率[21],且温度升高导致土壤有机碳加速分解[28]。本研究中樟子松林0—20 cm土层ER1和ER2较低,而ER3却高于其他3种林型,可能是由于活性碳组分对于环境的高敏感性,在更高活性中并不稳定,而樟子松林有机质含量较低,总有机碳趋于活化以供微生物和细根利用的碳源有限,无法支撑高活性有机碳的储存,因此,大多数以低活性有机碳的形式被利用,与樊蓉源等[29]的研究结果相近。
土壤碳库活度表明,4种林型各土层有机碳稳定性依次为兴安落叶松林>樟子松林>白桦林>山杨林,即针叶林土壤有机碳稳定性大于阔叶林,与前人[9]研究结论相近。这可能与2个方面有关:一是凋落物种类的影响[9, 24]。相关研究[24]表明,阔叶林叶片中淀粉、糖类等物质高于针叶林,这些物质中的碳大多数具有“不稳定性”,易被微生物分解利用后释放到土壤中,加之阔叶林凋落物较高的pH和N含量,导致其凋落物分解速率显著高于针叶林,使土壤固碳速率降低[27];而针叶凋落叶中较高的萜类物质和酚类物质易形成酸性腐殖质等衍生物[24],且针叶中的脂肪族化合物的烷基C比例和烷基C/O与烷基C的比例高于阔叶树种[27],在凋落物分解输入土壤后提高有机碳的稳定性。二是与土壤团聚体尺寸密切相关[6, 8]。朱家琪等[15]研究表明,该研究区阔叶林土壤中大团聚体含量高于针叶林。大团聚体可保护植物根系、微生物菌丝及其分泌物等高、中活性有机物质避免被氧化[6];而微团聚体中与矿物结合的多糖等低活性有机碳组分稳定,其碳周转率比大团聚体中的高、中活性有机物质慢[30]。LI等[31]研究发现,pH通过影响微生物活性及阳离子的凝聚和离子交换量,从而间接影响活性碳的含量和分布,且pH的增加导致土壤溶液中负电荷上升,使土壤溶解度增加,进而加剧土壤有机碳的损失。本研究中白桦林由于其较低的pH抑制微生物的生长活性,促进土壤碳库的积累,因此稳定性高于山杨林。
基于月尺度,本研究发现,针叶林各土层土壤有机碳稳定性于5月显著大于阔叶林,考虑与其凋落物分解速率有关。已有研究[24, 27]表明,土壤有机碳稳定性与凋落物性质密切相关。张頔等[32]在该研究区发现,由于5月土壤冻融交替作用导致凋落物受到热胀冷缩的机械力作用,而拥有硬质叶片的针叶抵抗这种机械力作用的能力远大于阔叶林,因此分解速率显著低于阔叶林[33]。同时,本研究发现,4种林型土壤碳库均在8月表现出不稳定状态,且阔叶林10—20 cm土层有机碳稳定性大于针叶林。是因为温度的升高刺激微生物代谢过程和蔗糖酶的活性[28]。已有研究[34]表明,蔗糖酶能够催化凋落物和土壤中的有机质并使其快速降解,其降解产物如葡萄糖和果糖等能够被微生物快速转化为能量和CO2,从而降低土壤有机碳的稳定性。但由于针叶林土壤中的有机碳大多以轻组有机碳的形式存在,在温度较高的情况下易被分解[10, 28],而阔叶林中的矿物结合碳浓度较高,因此导致阔叶林深层土壤有机碳稳定性高于针叶林,与SU等[10]在亚热带地区研究结论一致。兴安落叶松林于8—9月0—10 cm土层土壤有机碳稳定性显著低于樟子松林,考虑是由于其林下植被的多样性及凋落物种类的复杂性[3, 24],土壤微生物活性及养分释放速率加快[35],土壤固碳速率降低。由于本文中的影响因素仅涉及气候因子和土壤因子,并未测定凋落物层的相关因素,且该研究区中还分布着大量的针阔混交林,后续在此研究基础上可增加对混交林土壤碳库方面的研究,同时重视凋落物在土壤碳库循环中的作用,为全面衡量寒温带森林土壤碳库平衡和造林树种选择方面提供科学理论依据。
4. 结论
(1) 4种林型0—20 cm土层土壤LOC-H和LOC-M含量均于8月达到峰值,且阔叶林含量高于针叶林;LOC-H、LOC-M与TOC具有较好的相关性,均可用来预测土壤碳库变化,且LOC-H对于土壤碳库变化的响应更迅速。
(2) 土壤碳库中占主导地位的是氧化活性较低的稳定性有机碳,且针叶林土壤有机碳库整体稳定性大于阔叶林;但在较高温度下,阔叶林深层土壤固碳能力高于针叶林。
(3) 土壤活性有机碳含量和碳库稳定性受气候因素和土壤因素的共同作用。土壤蔗糖酶活性(SUC)、可溶性有机氮(DON)、降雨量(QR)和光合有效辐射(ESR)均为影响土壤活性碳含量和碳库稳定性的主导因子。建议在未来研究中增加对混交林和凋落物层在土壤碳库影响方面的研究。
-
图 1 不同森林类型土壤基本理化性质及酶活性变化特征
注:Y1表示左轴柱状图对应的数值,Y2表示右轴折线图对应的数值;图柱上方不同大写字母表示同一林型不同月份间差异达显著水平(p < 0.05);不同小写字母表示同一月份不同林型间差异达到显著水平(p < 0.05)。下同。
Fig. 1 Characteristics of changes in basic physical and chemical properties and enzyme activities of soils of different forest types
下载:
全尺寸图片
图 2 不同森林类型土壤TOC含量变化特征
Fig. 2 Changes of soil total organic carbon contents under different forest types
下载:
全尺寸图片
图 3 不同森林类型土壤易氧化碳组分含量变化特征
Fig. 3 Characteristics of changes in the contents of carbon oxidisable fractions in soils of different forest types
下载:
全尺寸图片
图 4 不同森林类型下土壤活性碳利用率变化特征
Fig. 4 Changes of soil active carbon utilization rates under different forest types
下载:
全尺寸图片
图 5 不同森林类型下土壤活性碳与影响因子冗余分析
注:TOC为土壤总有机碳;LOC-H为高活性易氧化有机碳;LOC-M为中活性易氧化有机碳;LOC-L为低活性易氧化有机碳;ER为活性有机碳利用率;CPA为土壤碳库活度;MT为月均温;RH为空气相对湿度;QR为每月总降雨量;ESR为太阳有效辐射;TN为全氮;SUC为蔗糖酶;MBN为微生物氮;DON为可溶性有机氮;NO3--N为硝态氮;NH4+-N为铵态氮;ST为土壤温度;SWC为土壤含水率;pH为土壤酸碱度。
Fig. 5 Redundancy analysis of soil active carbon and influencing factors under different forest types
下载:
全尺寸图片
表 1 研究区2023年5—9月相关气候因素
Table 1 Changes in relevant climatic factors in the study area, from May to September 2023
月份 空气温度/℃ 相对湿度/% 降雨量/mm 太阳有效辐射/(W·m-2) 5 9.26 54.87 51.40 107.95 6 15.66 73.52 59.10 131.11 7 20.17 75.86 37.70 135.37 8 16.89 70.71 33.70 112.63 9 8.98 70.07 26.20 56.42 注:表中空气湿度、相对湿度和太阳有效辐射均为当月平均值;降雨量为当月总降雨量。 表 2 样地基本情况
Table 2 Basic information of the sample sites
森林类型 海拔/m 林分密度/(株·hm-2) 坡度/(°) 郁闭度 平均胸径/cm 平均树高/m 树种组成 下木种类 LG 305 1 260 6 0.7 14.10±6.2 19.20±5.6 8落1樟1白 1,2,3,4 PS 290 736 10 0.6 21.36±4.9 22.37±3.8 8樟2落 1,3,5 BP 378 1 663 6 0.8 11.50±5.5 13.10±4.1 10白 1,3,4,6 PD 385 975 7 0.7 16.20±6.3 16.50±6.8 8杨2白 6,7,8 注:LG为兴安落叶松林;PS为樟子松林;BP为白桦林;PD为山杨林;DBH为胸径;表中数据为平均值±标准差;1为兴安杜鹃(Rhododen drondauricum);2为杜香(Ledum palustre);3为越桔(Vaccinium vitisidaea);4为笃斯越桔(Vaccinium uliginosum);5为羽节蕨(Gymnocarpium jessoense);6为红花鹿蹄草(Pyrola incarnata);7为舞鹤草(Maianthemum bifolium);8为东方草莓(Fragaria orientalis)。下同。 表 3 不同森林类型下土壤碳库活度变化特征
Table 3 Characteristics of changes in soil carbon pool activity under different forest types
森林类型 土层深度/cm 土壤碳库活度 5月 6月 7月 8月 9月 均值 LG 0-10 0.22±0.01Dc 0.21±0Dc 0.30±0.02Bb 0.35±0.01Ab 0.26±0.03Cb 0.27±0.05 10-20 0.22±0.01Bb 0.21±0Bab 0.29±0.03Aa 0.30±0.01Aa 0.20±0.01Bb 0.25±0.05 PS 0-10 0.30±0.01Ab 0.26±0.03Bb 0.31±0.02Aab 0.32±0.02Ac 0.22±0.01Bc 0.29±0.05 10-20 0.24±0.01Bb 0.22±0.01Cab 0.29±0.01Aa 0.29±0.01Aa 0.21±0.03Cb 0.25±0.04 BP 0-10 0.38±0.01Aa 0.22±0.01Cc 0.26±0.02Bc 0.37±0.01Aa 0.26±0.01Bb 0.30±0.07 10-20 0.32±0.06Aa 0.20±0.02Cb 0.24±0.03BCa 0.28±0.02ABa 0.24±0.02BCab 0.26±0.06 PD 0-10 0.38±0.01Aa 0.30±0.01Ba 0.36±0.01Ba 0.38±0.01Aa 0.31±0.01Ba 0.35±0.04 10-20 0.36±0.01Aa 0.23±0.01Ba 0.24±0.01Ba 0.25±0.01Bb 0.24±0.01Ba 0.27±0.05 注:表中数据为平均值±标准差;同列不同大写字母表示同一林型不同月份间差异达显著水平(p < 0.05);不同小写字母表示同一月份不同林型间差异达到显著水平(p < 0.05)。下同。 -
[1] 刘世荣, 王晖, 李海奎, 等. 碳中和目标下中国森林碳储量、碳汇变化预估与潜力提升途径[J]. 林业科学, 2024, 60(4): 157-172. LIU S R, WANG H, LI H K, et al. Projections of China's forest carbon storage and sequestration and ways of their potential capacity enhancement[J]. Scientia Silvae Sinicae, 2024, 60(4): 157-172. [2] 赵广, 张扬建. 大气CO2浓度升高对土壤碳库稳定性的影响[J]. 生态学报, 2023, 43(20): 8493-8503. ZHAO G, ZHANG Y J. Effect of elevated CO2 on the persistence of soil carbon pool[J]. Acta Ecologica Sinica, 2023, 43(20): 8493-8503. [3] 习丹, 翁浩东, 胡亚林, 等. 林冠氮添加和林下植被去除对杉木林土壤有机碳组分的影响[J]. 生态学报, 2021, 41(21): 8525-8534. XI D, WENG H D, HU Y L, et al. Effects of canopy nitrogen addition and understory removal on soil organic carbon fractions in a Chinese fir plantation[J]. Acta Ecologica Sinica, 2021, 41(21): 8525-8534. [4] 翟淑君, 列志旸, 卢嘉华, 等. 增温对南亚热带季风常绿阔叶林土壤有机碳及其组分的影响[J]. 生态学报, 2024, 44(13): 5508-5518. ZHAI S J, LIE Z Y, LU J H, et al. Effects of warming on soil organic carbon and its fractions of evergreen broad-leaved forests in southern subtropical monsoon region[J]. Acta Ecologica Sinica, 2024, 44(13): 5508-5518. [5] LOGINOW W, WISNIEWSKI W, GONET S, et al. Fractionation of organic carbon based on susceptibility to oxidation[J]. Polish Journal of Soil Science, 1987, 20(1): 47-52. [6] 李映雪, 臧真凤, 张瑜, 等. 退耕还草对土壤碳库活度及团聚体活性有机碳组分分布的影响[J]. 水土保持研究, 2023, 30(5): 241-249. LI Y X, ZANG Z F, ZHANG Y, et al. Changes in soil carbon pool activity and distribution of labile organic carbon composition in soil aggregates following conversion of farmland to grassland on the Loess Plateau[J]. Research of Soil and Water Conservation, 2023, 30(5): 241-249. [7] 姚旭, 景航, 梁楚涛, 等. 人工油松林表层土壤团聚体活性有机碳含量对短期氮添加的响应[J]. 生态学报, 2017, 37(20): 6724-6731. YAO X, JING H, LIANG C T, et al. Response of labile organic carbon content in surface soil aggregates to short-term nitrogen addition in artificial Pinus tabulaeformis forests[J]. Acta Ecologica Sinica, 2017, 37(20): 6724-6731. [8] WANG B, HAO S, ZHANG Q L. Protection mechanisms and influencing factors of soil organic carbon pools in the Larix gmelinii forests[J]. Ecological Indicators, 2023, 150: e110242. doi: 10.1016/j.ecolind.2023.110242 [9] DEVI A S. Influence of trees and associated variables on soil organic carbon: A review[J]. Journal of Ecology and Environment, 2021, 45(1): e5. [10] SU F L, XU S, SAYER E J, et al. Distinct storage mechanisms of soil organic carbon in coniferous forest and evergreen broadleaf forest in tropical China[J]. Journal of Environmental Management, 2021, 295: e113142. [11] 贾匡迪, 王勇强, 高雨, 等. 山西太岳山不同针叶纯林及混交林土壤有机碳库的变异性[J]. 东北林业大学学报, 2024, 52(3): 112-118. JIA K D, WANG Y Q, GAO Y, et al. Variability of soil organic carbon pool in different coniferous pure forests and mixed forests in Taiyue Mountain, Shanxi Province[J]. Journal of Northeast Forestry University, 2024, 52(3): 112-118. [12] SCHARLEMANN J P, TANNER E V, HIEDERER R, et al. Global soil carbon: Understanding and managing the largest terrestrial carbon pool[J]. Carbon Management, 2014, 5(1): 81-91. [13] 高明磊, 满秀玲, 段北星. 林下植被和凋落物对我国寒温带天然林土壤CO2通量的短期影响[J]. 北京林业大学学报, 2021, 43(3): 55-65. GAO M L, MAN X L, DUAN B X. Short-term effects of understory vegetation and litter on soil CO2 flux of natural forests in cold temperate zone of China[J]. Journal of Beijing Forestry University, 2021, 43(3): 55-65. [14] 魏红, 满秀玲. 中国寒温带不同林龄白桦林碳储量及分配特征[J]. 植物生态学报, 2019, 43(10): 843-852. WEI H, MAN X L. Carbon storage and its allocation in Betula platyphylla forests of different ages in cold temperate zone of China[J]. Chinese Journal of Plant Ecology, 2019, 43(10): 843-852. [15] 朱家琪, 满秀玲, 王飞. 我国寒温带四种森林植被类型下土壤团聚体粒级组成及其稳定性比较研究[J]. 土壤通报, 2020, 51(3): 606-613. ZHU J Q, MAN X L, WANG F. Composition and stability of soil aggregates in four forest vegetation types in cold temperate zone[J]. Chinese Journal of Soil Science, 2020, 51(3): 606-613. [16] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 1999: 455-456. LU R K. Soil agrochemical analytical methods[M]. Beijing: China Agricultural Science and Technology Press, 1999: 455-456. [17] QIAO L L, ZHOU H K, WANG Z H, et al. Variations in soil aggregate stability and organic carbon stability of alpine meadow and shrubland under long-term warming[J]. Catena, 2023, 222: e106848. [18] 梁彩群, 刘国彬, 王国梁, 等. 黄土高原人工刺槐林土壤团聚体中不同活性有机碳从南到北的变化特征[J]. 环境科学学报, 2020, 40(3): 1095-1102. LIANG C Q, LIU G B, WANG G L, et al. Variation characteristics of different labile organic carbon in soil aggregates of Robinia pseudoacacia plantation from south to north in the Loess Plateau[J]. Acta Scientiae Circumstantiae, 2020, 40(3): 1095-1102. [19] 王珍, 常顺利, 王冠正, 等. 天山中段北坡森林土壤有机碳库稳定性组分沿海拔的分异规律[J]. 生态学报, 2023, 43(18): 7390-7402. WANG Z, CHANG S L, WANG G Z, et al. Altitudinal distribution of the forest soil organic carbon fractions on the northern slope of the middle Tianshan Mountains[J]. Acta Ecologica Sinica, 2023, 43(18): 7390-7402. [20] MAKAROV M I, MALYSHEVA T I, MULYUKOVA O S, et al. Freeze-thaw effect on the processes of transformation of carbon and nitrogen compounds in alpine meadow soils[J]. Russian Journal of Ecology, 2015, 46(4): 317-324. [21] LIU Y, LI S Y, SUN X Y, et al. Variations of forest soil organic carbon and its influencing factors in East China[J]. Annals of Forest Science, 2016, 73(2): 501-511. [22] MIKI T, DOIH. Leaf phenological shifts and plant-microbe-soil interactions can determine forest productivity and nutrient cycling under climate change in an ecosystem model[J]. Ecological Research, 2016, 31(2): 263-274. [23] 李富, 臧淑英, 刘赢男, 等. 冻融作用对三江平原湿地土壤活性有机碳及酶活性的影响[J]. 生态学报, 2019, 39(21): 7938-7949. LI F, ZANG S Y, LIU Y N, et al. Effects of freezing and thawing on soil active organic carbon and enzyme activity in the Sanjiang Plain wetlands[J]. Acta Ecologica Sinica, 2019, 39(21): 7938-7949. [24] 陈子豪, 焦泽彬, 刘谣, 等. 凋落物季节性输入对川西亚高山森林土壤活性有机碳的影响[J]. 应用与环境生物学报, 2021, 27(3): 594-600. CHEN Z H, JIAO Z B, LIU Y, et al. Influences of seasonal litter input on soil active organic carbon in subalpine forests in western Sichuan[J]. Chinese Journal of Applied and Environmental Biology, 2021, 27(3): 594-600. [25] 辜翔, 张仕吉, 项文化, 等. 中亚热带4种森林类型土壤活性有机碳的季节动态特征[J]. 植物生态学报, 2016, 40(10): 1064-1076. GU X, ZHANG S J, XIANG W H, et al. Seasonal dynamics of active soil organic carbon in four subtropical forests in Southern China[J]. Chinese Journal of Plant Ecology, 2016, 40(10): 1064-1076. [26] 史进纳, 蒋代华, 肖斌, 等. 不同连栽代次桉树林土壤有机碳演变特征[J]. 热带作物学报, 2015, 36(4): 748-752. SHI J N, JIANG D H, XIAO B, et al. Evolution characteristics of soil organic carbon in different generation of continuous eucalyptus plantations[J]. Chinese Journal of Tropical Crops, 2015, 36(4): 748-752. [27] 高雨. 山西太岳山针叶林土壤有机碳稳定性变化特征及其影响因素分析[D]. 北京: 北京林业大学, 2021. GAO Y. Variability and influencing factors of soil organie carbon stability of coniferous forest in Taiyue Mountain, Shanxi Province[D]. Beijing: Beijing Forestry University, 2021. [28] JANZEN H H. Beyond carbon sequestration: Soil as conduit of solar energy[J]. European Journal of Soil Science, 2015, 66(1): 19-32. [29] 樊容源, 叶绍明, 张钱春, 等. 杉木纯林及其混交林土壤团聚体活性有机碳组分特征[J]. 西北林学院学报, 2023, 38(6): 20-28. FAN R Y, YE S M, ZHANG Q C, et al. Characteristics of active organic carbon components in soil aggregates of pure plantation of C. lanceolata and its mixed forest[J]. Journal of Northwest Forestry University, 2023, 38(6): 20-28. [30] SOUCÉMARIANADIN L N, CÉCILLON L, GUENET B, et al. Environmental factors controlling soil organic carbon stability in French forest soils[J]. Plant and Soil, 2018, 426(1): 267-286. [31] LI Y, WU H D, WANG J Z, et al. Plant biomass and soil organic carbon are main factors influencing dry-season ecosystem carbon rates in the coastal zone of the Yellow River Delta[J]. PLoS One, 2019, 14(1): e0210768. [32] 张頔, 满秀玲, 刘思琪, 等. 寒温带地区非生长季典型森林群落凋落物分解及养分释放[J]. 北京林业大学学报, 2022, 44(3): 65-74. ZHANG D, MAN X L, LIU S Q, et al. Litter decomposition and nutrient release of typical forest communities in non-growing season in cold temperate zone[J]. Journal of Beijing Forestry University, 2022, 44(3): 65-74. [33] VESTERDAL L, CLARKE N, SIGURDSSON B D, et al. Do tree species influence soil carbon stocks in temperate and boreal forests?[J]. Forest Ecology and Management, 2013, 309: 4-18. [34] 张晓曦, 田爽, 曾磊, 等. 北方常见绿化树种凋落物添加对城市土壤有机碳组成特征的影响[J]. 植物科学学报, 2024, 42(2): 140-149. ZHANG X X, TIAN S, ZENG L, et al. Effects of litter addition from common greening tree species in Northern China on organic carbon composition characteristics of urban soils[J]. Plant Science Journal, 2024, 42(2): 140-149. [35] 杨阳, 王宝荣, 窦艳星, 等. 植物源和微生物源土壤有机碳转化与稳定研究进展[J]. 应用生态学报, 2024, 35(1): 111-123. YANG Y, WANG B R, DOU Y X, et al. Advances in the research of transformation and stabilization of soil organic carbon from plant and microbe[J]. Chinese Journal of Applied Ecology, 2024, 35(1): 111-123.
