
Effects of Biological Crust Development on Soil Saturated Hydraulic Conductivity in the Three Gorges Reservoir Area
-
摘要:
目的 为了探究三峡库区生物结皮发育对土壤饱和导水率的影响及其驱动机制。 方法 在经果林下选取5个盖度梯度(0~20%, 20%~40%, 40%~60%, 60%~80%, 80%~100%)的苔藓结皮样地, 以林下无生物结皮覆盖样地为对照, 通过野外采样和室内分析测定不同盖度梯度生物结皮的土壤饱和导水率和基本土壤理化性质, 利用非线性回归、偏最小二乘回归和结构方程模型等方法明确经果林地土壤饱和导水率随生物结皮盖度的变化规律及其主控因素。 结果 (1) 经果林下生物结皮的发育显著增加土壤饱和导水率, 与对照相比, 0~20%, 20%~40%, 40%~60%, 60%~80%, 80%~100%生物结皮盖度梯度的土壤饱和导水率分别增加36.98%, 338.09%, 407.17%, 900.66%, 713.11%。(2)随着生物结皮盖度增加, 土壤饱和导水率呈近似对数函数趋势增加, 当生物结皮盖度>60%时, 土壤饱和导水率趋于稳定。(3)土壤饱和导水率与生物结皮特性及其驱动的土壤理化性质变化显著相关, 生物结皮厚度、土壤容重、总孔隙度、饱和含水率和水稳性团聚体含量是导致不同生物结皮盖度土壤饱和导水率存在差异的主控因素。(4)结构方程模型结果表明, 生物结皮的发育主要通过间接改良土壤结构(间接通径系数1.655>直接路径系数0.887)来增加土壤饱和导水率。 结论 研究结果为三峡库区经果林地生物结皮发育对水文和侵蚀过程的认识提供理论基础, 也为经果林地水土流失的防控提供实践指导。 Abstract:Objective Soil saturated hydraulic conductivity (Ks) reflects soil infiltration capacity and erosion resistance potential, which significantly affects hydrology and erosion processes. Economic fruit forest is an important land use type in the Three Gorges Reservoir area, biological crust wide-developed under the forest, which will inevitably drive the change in soil physical and chemical properties, thereby affecting Ks, but the effect of biological crust development on soil saturated hydraulic conductivity and its driving mechanism in reservoir area is still unclear. Methods Therefore, in order to investigate the effect of biological crust development on Ks in the Three Gorges Reservoir area. One site without biological crust (as control) and five moss crust sites with different coverage (0~20%, 20%~40%, 40%~60%, 60%~80%, 80%~100%) were selected under the economic fruit forest. The soil saturated hydraulic conductivity and basic soil physical and chemical properties of biological crust with different coverage gradients were determined by field sampling and laboratory analysis, combined with nonlinear regression, partial least squares regression and structural equation model methods. The changes in Ks with biological crust coverage and main factors attributed to these changes were clarified. Results (1) The development of biological crust significantly increased Ks, and compared to control, Ks of economic fruit forest covered by 0~20%, 20%~40%, 40%~60%, 60%~80%, 80%~100% biological crust increased by 36.98%, 338.09%, 407.17%, 900.66%, 713.11%, respectively. (2) Ks increased in an approximate logarithmic trend with the increase of biological crust coverage. When biological crust coverage was greater than 60%, Ks tended to be stable. (3) Ks was significantly correlated with the biological crust characteristics and the differences in soil physical and chemical properties driven by biological crust. The thickness of biological crust, soil bulk density, total porosity, saturated water content and water-stable aggregate content were the main influencing factors leading to the differences in Ks with different biological crust coverages. (4) The result of structural equation model showed that the development of biological crust was mainly through its indirect effect in improving soil structure (indirect path coefficient 1.655 bigger than direct path coefficient 0.887) to increase Ks. Conclusion These results provide a theoretical basis for understanding the hydrologic and erosion processes of biological crust development in the Three Gorges Reservoir area, and also provide practical guidance for the prevention and controlling of soil and water loss in the economic fruit forest land. -
土壤饱和导水率(soil saturated hydraulic conductivity, Ks)是指土壤在饱和状态,即土壤孔隙全部充满水时的土壤导水速率,是土壤容重、孔隙度和力学组成等多种物理特性的综合参数,其数值近似于土壤稳定入渗速率[1-3],反映土壤入渗能力和土壤抵抗侵蚀的潜力,对土壤侵蚀和水文过程具有重要影响。
生物土壤结皮(简称生物结皮)是由隐花植物如蓝藻、荒漠藻、地衣、苔藓类和土壤中微生物,以及相关的其他生物体通过菌丝体、假根和分泌物等与土壤表层颗粒胶结形成的十分复杂的复合体,其厚度通常为3~10 mm,有时甚至可达35 mm[4]。已有研究[5-8]表明,生物结皮的生长发育除直接影响饱和导水率外,也驱动土壤近地表理化性质发生变化,进而影响土壤饱和导水率,但目前关于生物结皮对土壤饱和导水率的影响仍存在分歧,大致可为促进与抑制。促进者[9-11]认为, 土壤生物结皮的形成不仅增加有机质含量,其假根及菌丝的存在促进团聚体的形成,增大土壤孔隙度,改善土壤结构,进而增强土壤饱和导水率;抑制者[12]认为, 生物结皮中的有机物遇水膨胀后,挤压作用导致土壤孔隙减小,从而降低饱和导水率。生物结皮的形成增强表层土壤的斥水性,进而抑制水分入渗,降低饱和导水率[13-14],且抑制作用随着生物结皮盖度的增大而增强[6, 15]。
三峡库区作为我国典型的生态脆弱区和水土流失重点治理区,近年来受三峡水库建设和库区移民活动的影响,库区农业产业结构和土地利用格局发生显著变化,主要表现为经果林面积持续增加,沿河流水系呈带状的扩张趋势,且主要分布在15°~25°的坡面上[16-17]。经果林下剧烈的人为扰动导致林下地表裸露,显著加剧水土流失,使得经果林逐渐成为三峡库区水土流失的重要策源地。而三峡库区良好的水热条件使得经果林下生物结皮广泛发育,作为陆地生态系统的重要组成部分,生物结皮的生长发育必然驱动土壤理化性质发生变化,进而影响土壤饱和导水率,但目前生物结皮发育对土壤饱和导水率的影响仍不清楚,因此,亟需开展相关研究,明确三峡库区经果林下生物结皮发育对土壤饱和导水率的影响及机制。虽然目前关于生物结皮对土壤饱和导水率的影响开展大量研究,并取得丰硕成果,但以往的研究主要集中在干旱及半干旱区,而生物结皮的生长发育受一系列生物与非生物因素的共同作用,其演替过程、群落组成及生态功能等呈现明显的过程复杂性和空间分异性。某一地区、特定土壤、特定生物结皮类型的研究结果并不能直接外推至其他地区。三峡库区的气候特征、土壤类型和地形等自然环境与其他地区存在明显差异,必然导致该地区土壤饱和导水率对生物结皮生长的响应也存在差异,但目前关于生物结皮的生长如何响应土壤饱和导水率及其响应机制仍不清楚。结合现阶段三峡库区经果林的快速扩张及严重水土流失和面源污染问题,科学剖析三峡库区经果林生物结皮生长对土壤饱和导水率的潜在影响,揭示其作用机制,对库区经果林水文和侵蚀过程的认识具有重要的意义,以期为库区经果林地水土流失和面源污染物的防控提供实践指导。
1. 材料与方法
1.1 研究区概况
研究区位于重庆市三峡库区忠县涂井乡友谊村的秦岭小流域(108°E,30°N),海拔为120~800 m,毗邻长江干流,属于典型的山地丘陵峡谷地带区。气候类型为典型的湿润季风气候,多年平均降水量为1 100 mm,多年平均气温为19 ℃,降雨主要集中在雨季4—10月,占全年降水总量的70%左右。主要土壤类型为紫色土,具有物理风化强烈、土层薄、土壤的保水抗旱和抗侵蚀能力差的特点,极易受降雨击溅和径流冲刷侵蚀影响,导致严重的水土流失问题。流域内主要以林地、果园和坡耕地为主,研究区内生物结皮主要分布在经果林下,林下生物结皮盖度为0~100%,但其盖度分布与经果林冠层盖度和林下草本盖度密切相关,通常经果林冠层盖度越大,林下草本相对较少时,生物结皮广泛发育。因此,一般经果林盖度>50%以上,林下草本较少时,生物结皮广泛发育,平均盖度能够达到50%以上,而经果林盖度较小时,林下草本盖度较高时,生物结皮发育相对较少,盖度相对较低。近年来受剧烈人类活动影响(松土、锄草、施肥等),导致林下地表裸露,水土流失加剧,使得经果林逐渐成为研究区水土流失的重要策源地,严重影响其生态安全。
1.2 样品采集与测定方法
2023年7月,在野外调查的基础上,在秦岭小流域的柑橘林下选取5种不同盖度等级(0~20%,20%~40%,40%~60%,60%~80%和80%~100%)的苔藓生物结皮样地,并以附近无结皮发育的柑橘林作为对照(CK)。各选取样地的柑橘林建设年限,冠层盖度、种植密度、坡位、坡向和坡度等基本一致,且空间上无重叠。生物结皮盖度采用照相法,将0.5 m×0.5 m的样方框放置在选定的监测样点,用单反相机从正面拍摄样方框的全景图,后期在photoshop软件中勾绘出样方框内的生物结皮,并计算其所占的面积即为生物结皮盖度。在各盖度等级的样地内,按“S”形法选择3个采样点,现场用游标卡尺(精度0.01 mm)测量生物结皮厚度。测量时在样点采集小块完整的生物结皮,从4个不同方向测量生物结皮厚度,平均值即为该块生物结皮的厚度,每个样点测量3块生物结皮厚度,平均值是该样点的生物结皮厚度。
测量完生物结皮厚度后,在每个样点选取未被扰动的土壤表面,用标准环刀(直径5.05 cm、高5 cm)采集3个原状土,用于测量土壤体积质量、总孔隙度、毛管孔隙度、田间持水率和饱和含水率等土壤性质[18],其平均值为该样点数据值,每个生物结皮盖度样地共9个重复;用大环刀(直径10.0 cm、高5.0 cm)采集3个原状土用于测量土壤饱和导水率,其平均值为该样点的饱和导水率,每个生物结皮盖度样地共9个重复;用铝盒采集原状土,带回实验室自然风干后采用湿筛法测量土壤水稳性团聚体含量[19]。其中饱和导水率的测量过程是,将大环刀带回实验室浸泡至饱和后,在浸泡饱和的环刀土样上固定1个大小相同的空环刀,利用马氏瓶向空环刀内供水并保持水头不变,同时用烧杯收集饱和土样的渗水量,每隔5 min测量1次直至连续5次的渗水量相同,此时单位时间渗水量即为土壤饱和导水率,由于测量时水温保持一致(均为28 ℃),故未进行温度换算,计算公式为:
$$ K_{\mathrm{s}}=\frac{10 Q}{A t}\left(\frac{S}{S+H}\right) $$ (1) 式中:Ks为饱和导水率(mm/min);Q为渗水量(mL);A为土样的横截面积(cm2);t为渗水时间(min);S为土样高度(cm);H为水头高度(cm);10为单位换算系数。
采集完原状土后,在每个样地的3个样点分别采集一定量的表层(0—5 cm)散土样,然后将3个样点的散土样充分混合后取约1 kg带回实验室,自然风干后用于测定土壤的机械组成和有机碳质量分数。其中土壤机械组成采用马尔文激光粒度仪(Mastersizer 2000)测定;土壤有机碳质量分数采用硫酸—重铬酸钾氧化外加热法测定,每种土样重复3次。
1.3 数据处理
不同生物结皮盖度间生物结皮厚度、土壤理化性质和土壤饱和导水率的差异采用单因素方差分析(One-way ANOVA)中的最小显著差异分析(LSD)方法。生物结皮盖度与土壤饱和导水率之间的关系采用非线性回归方法确定。不同生物结皮盖度土壤饱和导水率差异的主要影响因素采用偏最小二乘回归(PLSR)方法进行确定,采用结构方程模型对土壤饱和导水率的主要影响因素进行路径分析。PLSR分析用SIMCA 14.1软件完成,结构方程模型采用Rstudio软件完成,其他数据分析采用SPSS 22.0软件完成,图制作采用Origin 2019b软件完成。
2. 结果与分析
2.1 生物结皮发育对土壤理化性质的影响
经果林下不同盖度等级生物结皮样地的生物结皮厚度和土壤理化性质的单因素方差分析结果见表 1。不同盖度生物结皮厚度为2.05~4.92 mm,表现为随生物结皮盖度增大呈先增大后减小的趋势,其中最大值出现在盖度等级为60%~80%,且显著大于其他盖度生物结皮厚度,而盖度等级为0~20%生物结皮厚度则显著小于其他盖度等级生物结皮厚度(p < 0.05)。
表 1 不同生物结皮盖度土壤理化性质的差异Table 1 Differences in soil physical and chemical properties with different biocrust coverage土壤理化性质 CK 0~20% 20%~40% 40%~60% 60%~80% 80%~100% 生物结皮厚度/mm — 2.05±0.28c 3.18±0.17b 3.67±0.69b 4.92±0.55a 3.65±0.75b 砂粒/% 23.92±0.48a 14.79±0.12b 13.15±0.52c 7.08±0.30f 11.20±0.12d 8.82±0.30e 粉粒/% 73.81±0.46f 82.33±0.11e 83.70±0.49d 89.21±0.29a 85.45±0.12c 87.78±0.28b 黏粒/% 2.27±0.03f 2.88±0.01e 3.15±0.02d 3.72±0.02a 3.35±0.01c 3.39±0.02b 土壤体积质量/(g·cm-3) 1.44±0.03a 1.39±0.08a 1.35±0.17a 1.31±0.16a 1.31±0.02a 1.30±0.07a 饱和含水率/% 29.96±1.99a 31.81±4.55a 35.48±9.92a 35.65±7.44a 35.67±1.40a 37.03±5.55a 田间持水量/% 25.27±1.64a 24.44±1.37a 31.98±9.55a 30.79±4.00a 27.25±1.82a 29.07±2.60a 总孔隙度/% 45.65±1.26a 47.32±3.10a 49.12±6.41a 50.50±6.23a 50.48±0.46a 50.93±2.36a 毛管孔隙度/% 41.01±0.81a 39.13±0.60a 44.06±5.49a 43.64±1.64a 41.52±1.43a 42.62±1.39a 有机质/(g·kg-1) 15.31±3.56b 22.52±0.93a 22.50±0.65a 22.97±0.02a 23.91±0.33a 23.51±1.13a 水稳性团聚体/% 54.60±0.43b 58.92±1.62a 59.74±1.13a 60.14±0.44a 60.96±1.25a 60.75±0.79a 注:表中数据为平均值±标准差;水稳性团聚体为>0.25 mm团聚体;同行不同字母表示不同处理间差异显著(p < 0.05)。 生物结皮的发育也显著影响土壤理化性质,且不同盖度等级间存在一定差异。与林下无生物结皮发育的样地相比,经果林下生物结皮的发育后土壤砂粒平均值显著减小53.99%,而粉粒和黏粒的平均值分别显著增大16.10%和45.32%,土壤颗粒表现为细化趋势。
经果林下生物结皮的发育也显著增加土壤有机质和水稳性团聚体,但不显著影响土壤体积质量、饱和体积分数、田间持水量、总孔隙度和毛管孔隙度。而随着生物结皮盖度的增加,砂粒表现为波动减小趋势,而粉粒和黏粒表现为波动增大趋势,且不同盖度间存在显著差异。随生物结皮盖度的增加土壤体积质量呈减小趋势,饱和体积分数、田间持水量、总孔隙度、毛管孔隙度、土壤有机质和水稳性团聚体均呈波动增大趋势,但不同盖度间没有显著差异。
2.2 生物结皮发育对土壤饱和导水率的影响
描述性统计结果表明,不同生物结皮盖度的最小饱和导水率值为0.22~3.25 mm/min,最大饱和导水率为0.44~3.66 mm/min,平均值为0.35~3.47 mm/min,变异系数为0.04~0.50,均<1.0,表现为中等变异性(表 2)。而经果林下生物结皮的发育显著增加土壤饱和导水率,与无生物结皮覆盖的经果林相比,0~20%,20%~40%,40%~60%,60%~80%,80%~100%生物结皮盖度梯度的土壤饱和导水率分别增加36.98%,338.09%,407.17%,900.66%,713.11%,除0~20%生物结皮盖度梯度的土壤饱和导水率表现为不显著增加外,其余生物结皮盖度梯度的土壤饱和导水率均表现为显著增加趋势。随着生物结皮盖度增加,土壤饱和导水率呈近似对数函数的趋势增加(图 1),当生物结皮盖度>60%,土壤饱和导水率趋于稳定。
表 2 不同生物结皮盖度土壤饱和导水率特征值Table 2 Characteristic values of soil saturated hydraulic conductivity with different biological crust coverage生物结皮盖度/% 最小值/(mm·min-1) 最大值/(mm·min-1) 平均值/(mm·min-1) 标准差 变异系数 CK 0.22 0.44 0.35 0.09 0.26 0~20 0.22 0.79 0.48 0.24 0.50 20~40 1.44 1.61 1.52 0.07 0.05 40~60 1.60 1.86 1.76 0.12 0.07 60~80 3.25 3.66 3.47 0.17 0.05 80~100 2.68 2.92 2.82 0.10 0.04 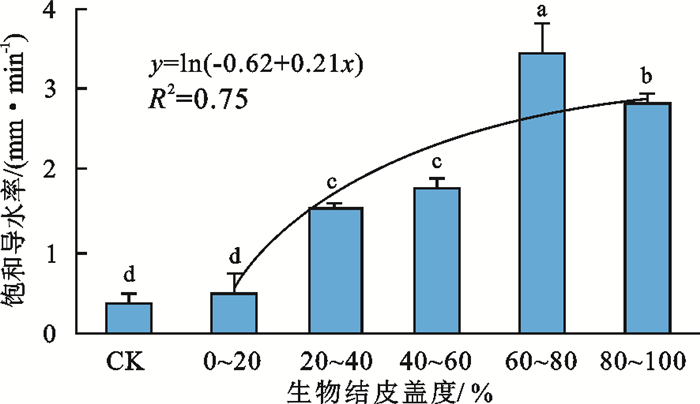 图 1 不同生物结皮盖度土壤饱和导水率的差异及变化规律Fig. 1 Differences and variation rules of saturated hydraulic conductivity of soils with different biological crust coverage
图 1 不同生物结皮盖度土壤饱和导水率的差异及变化规律Fig. 1 Differences and variation rules of saturated hydraulic conductivity of soils with different biological crust coverage 下载:
全尺寸图片
下载:
全尺寸图片
2.3 土壤饱和导水率的影响因素
采用Spearman相关分析法分析生物结皮厚度和土壤理化性质与土壤饱和导水率的相关性,结果表明,土壤饱和导水率随生物结皮厚度、饱和含水率、总孔隙度、有机质和水稳性团聚体的增加呈显著的线性增大趋势,而随土壤体积质量的增大呈显著的线性减小趋势(图 2)。采用偏最小二乘回归分析方法表明,测定的影响土壤饱和导水率的所有因素中,仅生物结皮厚度、土壤体积质量、总孔隙度、饱和含水率和水稳性团聚体的重要性投影(VIP)值>1,分别为1.22,1.20,1.20,1.15,1.07(图 3)。表明不同生物结皮盖度梯度经果林地土壤饱和导水率的差异主要受生物结皮厚度、土壤体积质量、总孔隙度、饱和含水率和水稳性团聚体的影响。
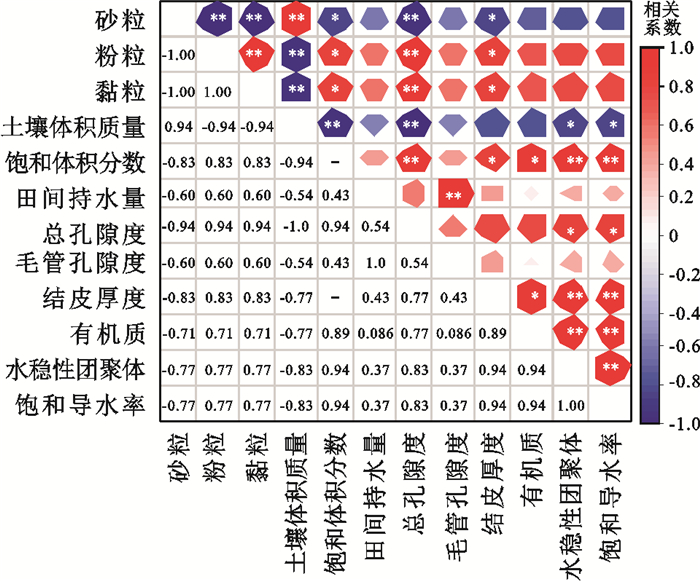 图 2 土壤饱和导水率、生物结皮厚度及土壤理化性质指标相关矩阵Fig. 2 Correlation matrix of soil saturated hydraulic conductivity, biological crust thickness and soil physical and chemical properties下载:
全尺寸图片
图 2 土壤饱和导水率、生物结皮厚度及土壤理化性质指标相关矩阵Fig. 2 Correlation matrix of soil saturated hydraulic conductivity, biological crust thickness and soil physical and chemical properties下载:
全尺寸图片
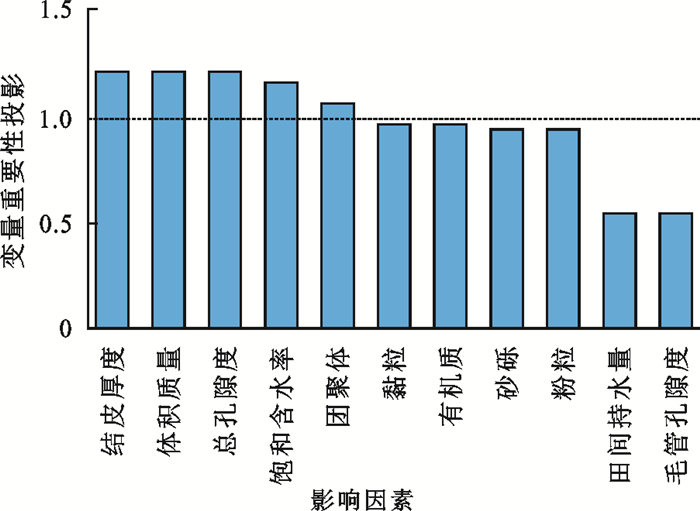 图 3 土壤饱和导水率各影响因素的变量重要性值Fig. 3 Variable importance values of each influencing factor of soil saturated hydraulic conductivity下载:
全尺寸图片
图 3 土壤饱和导水率各影响因素的变量重要性值Fig. 3 Variable importance values of each influencing factor of soil saturated hydraulic conductivity下载:
全尺寸图片
不同影响因素存在相互作用,为了进一步探究各主要影响因素对土壤饱和导水率的影响路径,采用结构方程模型进一步明确各主要影响因素对土壤饱和导水率的直接和间接效应。结果表明,土壤体积质量、水稳性团聚体、孔隙特征和生物结皮厚度的直接路径系数分别为-0.545,0.561,0.679和0.887,表明土壤体积质量的直接作用与土壤饱和导水率存在显著的负效应,而土壤水稳性团聚体、孔隙特征和生物结皮厚度的直接作用存在显著的正效应(图 4)。
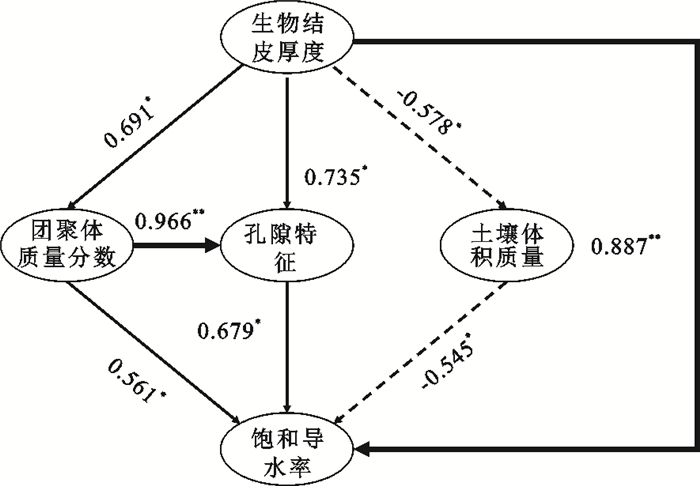 图 4 土壤饱和导水率与其主要影响因素的作用路径χ2/df=0.206;p=0.827;GFI=0.996;AGFI=0.967;RMSEA=0
图 4 土壤饱和导水率与其主要影响因素的作用路径χ2/df=0.206;p=0.827;GFI=0.996;AGFI=0.967;RMSEA=0
注:*表示0.05显著性水平下效应显著;**表示0.01显著性水平下效应显著;χ2为卡方,df为自由度,GFI为拟合优度指数,AGFI为调整后的拟合优度指数,RMSEA为近似均方根误差。Fig. 4 Action path of soil saturated water conductivity and its main influencing factors下载:
全尺寸图片
生物结皮除存在直接正效应外,生物结皮的发育也通过减小土壤体积质量、增加土壤水稳性团聚体、改善土壤结构进而间接影响土壤饱和导水率,其间接路径系数为1.655,大于其直接路径系数0.887。表明经果林下生物结皮的发育主要通过改善土壤结构来增大土壤饱和导水率。
3. 讨论
3.1 生物结皮发育对土壤饱和导水率的影响
生物结皮除自身的生长直接影响土壤饱和导水率外,也驱动土壤理化性质发生变化,进而影响土壤饱和导水率。本研究得出,有生物结皮覆盖经果林地的土壤饱和导水率明显高于无生物结皮覆盖的裸地,与KAKEH等[20]在伊朗黄土区、张思琪等[10]在喀斯特地貌和张冠华等[13]在三峡库区研究结论一致。本研究得出,土壤饱和导水率随着生物结皮盖度增加呈近似对数函数的趋势增加,当生物结皮盖度>60%时,土壤饱和导水率趋于稳定,与张思琪等[10]在喀斯特的研究中得出土壤入渗速率随生物结皮盖度的增大而增大的规律相同。其主要原因是苔藓结皮的假根浓密,使得样地柔软、疏松,其强大的吸附力黏结大量的土壤颗粒形成稳定的团聚结构,显著增加土壤孔隙度,从而提高土壤饱和导水率[13, 21-22]。本研究发现,土壤饱和导水率在生物结皮盖度为60%~80%时最大,而不是80%~100%盖度梯度的生物结皮样地土壤饱和导水率最大。可能是因为虽然生物结皮盖度大,但其蒸散量也大,受季节性干旱的影响,盖度越大的蒸散发导致的土壤水分损耗增大,进而可能抑制生物结皮的生长,而本研究也发现,80%~100%盖度梯度下生物结皮厚度小于60%~80%盖度下的生物结皮厚度(表 1)。野外条件下,80%~100%盖度梯度下生物结皮样地存在较大的土壤裂隙,也可能是大量水分蒸发导致的,这也间接证明高盖度条件下可能导致土壤水分的大量蒸发,进而抑制生物结皮的发育。但干旱、半干旱区的多数研究[8, 17, 23]均发现,土壤饱和导水率随生物结皮(藻结皮为主)盖度的增加呈现减小趋势,其主要与生物结皮类型有关。因为干旱、半干旱区生物结皮发育过程中,最先发育的为藻结皮,随着生物结皮的生长和演替,慢慢形成以藻结皮为主的藻-藓混合结皮,最后发育成以藓结皮为主的藓—藻混合结皮[6]。而藻结皮的发育形成一层致密的不透水层,使其表层结构更封闭,从而减小土壤孔隙,进而减小土壤饱和导水率[24]。而孙福海等[25]在黄土高原的研究也发现,随着生物结皮由藻向藓的发育演替,其土壤饱和导水率呈现先降低后增加的趋势,因此,生物结皮类型是影响土壤饱和导水率的重要因素。但李雨晨等[26]在气候条件和生物结皮类型与三峡库区相似的红壤区的研究发现,土壤饱和导水率随生物结皮盖度的增大而减小,这种差异主要与土壤类型有关。红壤为黏壤土,土壤黏粒质量分数较高,超过30%[27],而黏粒具有胀缩性,吸水后膨胀导致水分通道减少,从而降低土壤饱和导水率。而本研究土壤为砂壤土,砂粒质量分数较高,黏粒质量分数较少,少量的黏粒促进团聚体的发育,进而改善土壤结构,因此,本研究中土壤饱和导水随黏粒质量分数的增加而增加(图 2)。
3.2 不同盖度生物结皮土壤饱和导水率差异的影响机制
本研究通过变量重要性投影和结构方程模型得到主要影响因素的路径系数发现,生物结皮厚度对土壤饱和导水率的直接影响系数最大,且为正相关。是因为随着生物结皮生长发育,其苔藓、藻类等自养生物进行光合作用和细胞分泌,增强土壤动物和微生物活性。生物结皮本身积累的大量假根和菌丝在死亡之后分解,增加土壤有机质质量分数[28-29]。随着生物结皮生长发育,结皮层土壤的有机质质量分数随之增多,从而促进土壤团聚体的发育形成,提高水稳性团聚体质量分数,改善土壤孔隙度,减小土壤体积质量,进而提高土壤饱和导水率[30-31]。本研究中,生物结皮的发育主要通过间接作用来影响土壤饱和导水率,如减小土壤体积质量,增加土壤水稳性团聚体,改善土壤结构。并且其间接路径系数大于其直接路径系数,表明经果林下生物结皮的发育主要通过改善土壤结构的间接作用增大土壤饱和导水率。
多数干旱、半干旱区的研究[32-33]结果发现,生物结皮对土壤饱和导水率的影响主要受土壤性质的影响,与本研究的结论一致,生物结皮通过间接影响土壤性质来影响土壤饱和导水率。但大多黄土区研究[34-36]表明,土壤质地是影响土壤饱和导水率的主要因素,而本研究中生物结皮主要通过改善土壤结构的间接作用来增大土壤饱和导水率,二者产生差异的原因可能与生物结皮类型和区域气候及土壤特性有关。综上可知,生物结皮的发育通过改善土壤结构增加土壤饱和导水率,促进地表径流的入渗来增加水分的垂直入渗,减小地表径流量,进而减小地表径流的冲刷侵蚀[37]。生物结皮的发育也通过增加地表随机粗糙度来增加地表径流的入渗时间,同时增加径流的阻力系数,进而减小径流量和动能来影响坡面水文和侵蚀过程[10]。因此,生物结皮的发育通过调控降雨的入渗和产流过程来影响水文过程进而影响侵蚀过程。而面源污染物作为三峡库区另一重要生态环境问题,生物结皮发育驱动的水文和侵蚀过程的变化必然引起面源污染物在垂直和水平方向上的迁移过程也发生响应,但目前仍不清晰,未来需要进一步探究。
4. 结论
(1) 经果林下生物结皮的发育显著增加土壤饱和导水率,与无结皮发育样地相比,0~20%,20%~40%,40%~60%,60%~80%,80%~100%生物结皮盖度梯度的土壤饱和导水率分别增加36.98%,338.09%,407.17%,900.66%,713.11%。
(2) 随着生物结皮盖度增加,土壤饱和导水率呈近似对数函数的趋势增加,当生物结皮盖度>60%时,土壤饱和导水率趋于稳定。
(3) 土壤饱和导水率与生物结皮特性及其驱动的土壤理化性质变化显著相关,生物结皮厚度、土壤体积质量、总孔隙度、饱和含水率和水稳性团聚体是导致不同生物结皮盖度土壤饱和导水率存在差异的主控因素。
(4) 结构方程模型结果表明,生物结皮的发育主要通过间接改良土壤结构(间接通径系数1.655>直接路径系数0.887)来增加土壤饱和导水率。
-
图 1 不同生物结皮盖度土壤饱和导水率的差异及变化规律
Fig. 1 Differences and variation rules of saturated hydraulic conductivity of soils with different biological crust coverage
下载:
全尺寸图片
图 2 土壤饱和导水率、生物结皮厚度及土壤理化性质指标相关矩阵
Fig. 2 Correlation matrix of soil saturated hydraulic conductivity, biological crust thickness and soil physical and chemical properties
下载:
全尺寸图片
图 3 土壤饱和导水率各影响因素的变量重要性值
Fig. 3 Variable importance values of each influencing factor of soil saturated hydraulic conductivity
下载:
全尺寸图片
图 4 土壤饱和导水率与其主要影响因素的作用路径
χ2/df=0.206;p=0.827;GFI=0.996;AGFI=0.967;RMSEA=0
注:*表示0.05显著性水平下效应显著;**表示0.01显著性水平下效应显著;χ2为卡方,df为自由度,GFI为拟合优度指数,AGFI为调整后的拟合优度指数,RMSEA为近似均方根误差。Fig. 4 Action path of soil saturated water conductivity and its main influencing factors
下载:
全尺寸图片
表 1 不同生物结皮盖度土壤理化性质的差异
Table 1 Differences in soil physical and chemical properties with different biocrust coverage
土壤理化性质 CK 0~20% 20%~40% 40%~60% 60%~80% 80%~100% 生物结皮厚度/mm — 2.05±0.28c 3.18±0.17b 3.67±0.69b 4.92±0.55a 3.65±0.75b 砂粒/% 23.92±0.48a 14.79±0.12b 13.15±0.52c 7.08±0.30f 11.20±0.12d 8.82±0.30e 粉粒/% 73.81±0.46f 82.33±0.11e 83.70±0.49d 89.21±0.29a 85.45±0.12c 87.78±0.28b 黏粒/% 2.27±0.03f 2.88±0.01e 3.15±0.02d 3.72±0.02a 3.35±0.01c 3.39±0.02b 土壤体积质量/(g·cm-3) 1.44±0.03a 1.39±0.08a 1.35±0.17a 1.31±0.16a 1.31±0.02a 1.30±0.07a 饱和含水率/% 29.96±1.99a 31.81±4.55a 35.48±9.92a 35.65±7.44a 35.67±1.40a 37.03±5.55a 田间持水量/% 25.27±1.64a 24.44±1.37a 31.98±9.55a 30.79±4.00a 27.25±1.82a 29.07±2.60a 总孔隙度/% 45.65±1.26a 47.32±3.10a 49.12±6.41a 50.50±6.23a 50.48±0.46a 50.93±2.36a 毛管孔隙度/% 41.01±0.81a 39.13±0.60a 44.06±5.49a 43.64±1.64a 41.52±1.43a 42.62±1.39a 有机质/(g·kg-1) 15.31±3.56b 22.52±0.93a 22.50±0.65a 22.97±0.02a 23.91±0.33a 23.51±1.13a 水稳性团聚体/% 54.60±0.43b 58.92±1.62a 59.74±1.13a 60.14±0.44a 60.96±1.25a 60.75±0.79a 注:表中数据为平均值±标准差;水稳性团聚体为>0.25 mm团聚体;同行不同字母表示不同处理间差异显著(p < 0.05)。 表 2 不同生物结皮盖度土壤饱和导水率特征值
Table 2 Characteristic values of soil saturated hydraulic conductivity with different biological crust coverage
生物结皮盖度/% 最小值/(mm·min-1) 最大值/(mm·min-1) 平均值/(mm·min-1) 标准差 变异系数 CK 0.22 0.44 0.35 0.09 0.26 0~20 0.22 0.79 0.48 0.24 0.50 20~40 1.44 1.61 1.52 0.07 0.05 40~60 1.60 1.86 1.76 0.12 0.07 60~80 3.25 3.66 3.47 0.17 0.05 80~100 2.68 2.92 2.82 0.10 0.04 -
[1] WANG H, ZHANG G H, LI N N, et al. Soil erodibility as impacted by vegetation restoration strategies on the Loess Plateau of China[J]. Earth Surface Processes and Landforms, 2019, 44(3): 796-807. doi: 10.1002/esp.4531 [2] 肖波, 赵允格, 邵明安. 陕北水蚀风蚀交错区两种生物结皮对土壤饱和导水率的影响[J]. 农业工程学报, 2007, 23(12): 35-40. XIAO B, ZHAO Y G, SHAO M A. Effects of biological soil crust on saturated hydraulic conductivity in water-wind erosion crisscross region, North of Shaanxi Province, China[J]. Transactions of the Chinese Society of Agricultural Engineering, 2007, 23(12): 35-40. [3] LI Y Y, SHAO M A. Change of soil physical properties under long-term natural vegetation restoration in the Loess Plateau of China[J]. Journal of Arid Environments, 2006, 64(1): 77-96. doi: 10.1016/j.jaridenv.2005.04.005 [4] 李新荣, 贾玉奎, 龙利群, 等. 干旱半干旱地区土壤微生物结皮的生态学意义及若干研究进展[J]. 中国沙漠, 2001, 21(1): 4-11. LI X R, JIA Y K, LONG L Q, et al. Advances in microbiotic soil crust research and its ecological significance in arid and semiarid regions[J]. Journal of Desert Research, 2001, 21(1): 4-11. [5] 张光辉. 退耕驱动的近地表特性变化对土壤侵蚀的潜在影响[J]. 中国水土保持科学, 2017, 15(4): 143-154. ZHANG G H. Potential effects of changes in near soil surface characteristics driven by farmland abandonment on soil erosion[J]. Science of Soil and Water Conservation, 2017, 15(4): 143-154. [6] WANG H, ZHANG G H, LIU F, et al. Effects of biological crust coverage on soil hydraulic properties for the Loess Plateau of China[J]. Hydrological Processes, 2017, 31(19): 3396-3406. doi: 10.1002/hyp.11263 [7] DAN C X, LIU G, ZHAO Y G, et al. The effects of typical grass cover combined with biocrusts on slope hydrology and soil erosion during rainstorms on the Loess Plateau of China: An experimental study[J]. Hydrological Processes, 2023, 37(1): e14794. doi: 10.1002/hyp.14794 [8] SUN F H, XIAO B, KIDRON G J. Towards the influences of three types of biocrusts on soil water in drylands: Insights from horizontal infiltration and soil water retention[J]. Geoderma, 2022, 428: e116136. doi: 10.1016/j.geoderma.2022.116136 [9] ELDRIDGE D J. Cryptogams, vascular plants, and soil hydrological relations: Some preliminary results from the semiarid woodlands of eastern Australia[J]. The Great Basin Naturalist, 1993, 53(1): 48-58. [10] 张思琪, 张科利, 曹梓豪, 等. 喀斯特坡面生物结皮发育特征及其对土壤水分入渗的影响[J]. 应用生态学报, 2021, 32(8): 2875-2885. ZHANG S Q, ZHANG K L, CAO Z H, et al. Developmental characteristics of biological soil crusts and their effects on soil water infiltration on Karst slope[J]. Chinese Journal of Applied Ecology, 2021, 32(8): 2875-2885. [11] 张冠华, 易亮, 丁文峰, 等. 三峡库区苔藓生物结皮对土壤水分入渗的影响[J]. 应用生态学报, 2022, 33(7): 1835-1842. ZHANG G H, YI L, DING W F, et al. Effects of moss biocrust on soil water infiltration in the Three Gorges Reservoir Area, China[J]. Chinese Journal of Applied Ecology, 2022, 33(7): 1835-1842. [12] YANG H T, LIU L C, LI X R, et al. Characteristics of soil water repellency after sand dune stabilization in the Tengger Desert[J]. Sciences in Cold and Arid Regions, 2012, 4(5): 408-416. doi: 10.3724/SP.J.1226.2012.00408 [13] 张培培, 赵允格, 王媛, 等. 黄土高原丘陵区生物结皮土壤的斥水性[J]. 应用生态学报, 2014, 25(3): 657-663. ZHANG P P, ZHAO Y G, WANG Y, et al. Impact of biological soil crusts on soil water repellence in the hilly Loess Plateau Region, China[J]. Chinese Journal of Applied Ecology, 2014, 25(3): 657-663. [14] FISCHER T, VESTE M, WIEHE W, et al. Water repellency and pore clogging at early successional stages of microbiotic crusts on inland dunes, Brandenburg, NE Germany[J]. Catena, 2010, 80(1): 47-52. doi: 10.1016/j.catena.2009.08.009 [15] XIAO B, SUN F H, HU K L, et al. Biocrusts reduce surface soil infiltrability and impede soil water infiltration under tension and ponding conditions in dryland ecosystem[J]. Journal of Hydrology, 2019, 568: 792-802. doi: 10.1016/j.jhydrol.2018.11.051 [16] 刘彦随, 璩路路. 近30年三峡库区用地格局变化与人地系统演化[J]. 长江流域资源与环境, 2022, 31(8): 1664-1676. LIU Y S, QU L L. Land use pattern change and human-earth system evolution in Three Gorges Reservoir Area in recent 30 years[J]. Resources and Environment in the Yangtze Basin, 2022, 31(8): 1664-1676. [17] 王斌, 何丙辉, 林娜, 等. 奉节县脐橙果园用地时空演变特征与驱动因子分析[J]. 农业机械学报, 2021, 52(6): 204-214. WANG B, HE B H, LIN N, et al. Spatial and temporal evolution of navel orange orchard land and its driving factors in Fengjie County[J]. Transactions of the Chinese Society for Agricultural Machinery, 2021, 52(6): 204-214. [18] ZHU P Z, ZHANG G H, ZHANG B J. Soil saturated hydraulic conductivity of typical revegetated plants on steep gully slopes of Chinese Loess Plateau[J]. Geoderma, 2022, 412: e115717. [19] ZHU P Z, ZHANG G H, WANG H X, et al. Land surface roughness affected by vegetation restoration age and types on the Loess Plateau of China[J]. Geoderma, 2020, 366: e114240. [20] KAKEH J, GORJI M, MOHAMMADI M H, et al. Biocrust islands enhance infiltration, and reduce runoff and sediment yield on a heavily salinized dryland soil[J]. Geoderma, 2021, 404: e115329. [21] 程才, 李玉杰, 龙明忠, 等. 苔藓结皮在我国喀斯特石漠化治理中的应用潜力[J]. 应用生态学报, 2019, 30(7): 2501-2510. CHENG C, LI Y J, LONG M Z, et al. Application potential of bryophyte soil crust on the control of karst rocky desertification[J]. Chinese Journal of Applied Ecology, 2019, 30(7): 2501-2510. [22] 李冰, 张朝晖. 喀斯特石漠结皮层藓类物种多样性及在石漠化治理中的作用研究[J]. 中国岩溶, 2009, 28(1): 55-60. LI B, ZHANG Z H. Species diversity of mosses crust and the effect in karst rocky desertification control[J]. Carsologica Sinica, 2009, 28(1): 55-60. [23] 李宁宁, 张光辉, 王浩, 等. 黄土丘陵沟壑区生物结皮对土壤抗蚀性能的影响[J]. 中国水土保持科学, 2020, 18(1): 42-48. LI N N, ZHANG G H, WANG H, et al. Soil anti-erodibility influenced by biological crusts in Loess Hilly and Gully Region[J]. Science of Soil and Water Conservation, 2020, 18(1): 42-48. [24] 李新荣, 谭会娟, 回嵘, 等. 中国荒漠与沙地生物土壤结皮研究[J]. 科学通报, 2018, 63(23): 2320-2334. LI X R, TAN H J, HUI R, et al. Researches in biological soil crust of China: A review[J]. Chinese Science Bulletin, 2018, 63(23): 2320-2334. [25] 孙福海, 肖波, 李胜龙, 等. 黄土高原不同发育阶段生物结皮的导水和持水特征[J]. 草业学报, 2021, 30(6): 54-63. SUN F H, XIAO B, LI S L, et al. Effects of biological soil crusts in different developmental stages on soil water permeability and water holding capacity in the Chinese Loess Plateau[J]. Acta Prataculturae Sinica, 2021, 30(6): 54-63. [26] 李雨晨, 平原, 澹腾辉, 等. 红壤丘陵区不同盖度生物结皮对水分入渗的影响[J]. 水土保持学报, 2023, 37(5): 71-77. http://stbcxb.alljournal.com.cn/stbcxb/article/abstract/20230509?st=search LI Y C, PING Y, TAN T H, et al. Effects of biocrust with different coverage on water infiltration in red soil hilly region[J]. Journal of Soil and Water Conservation, 2023, 37(5): 71-77. http://stbcxb.alljournal.com.cn/stbcxb/article/abstract/20230509?st=search [27] 王冬冬, 高磊, 陈效民, 等. 红壤丘陵区坡地土壤颗粒组成的空间分布特征研究[J]. 土壤, 2016, 48(2): 361-367. WANG D D, GAO L, CHEN X M, et al. Spatial distribution characteristics of soil particle composition of slope land red soil region, China[J]. Soils, 2016, 48(2): 361-367. [28] CHAMIZO S, CANTÓN Y, MIRALLES I, et al. Biological soil crust development affects physicochemical characteristics of soil surface in semiarid ecosystems[J]. Soil Biology and Biochemistry, 2012, 49: 96-105. [29] GAO L Q, BOWKER M A, XU M X, et al. Biological soil crusts decrease erodibility by modifying inherent soil properties on the Loess Plateau, China[J]. Soil Biology and Biochemistry, 2017, 105: 49-58. [30] 高婵婵, 赵传燕, 王超, 等. 黑河上游天老池流域不同植被下土壤理化性质和入渗特征[J]. 水土保持学报, 2016, 30(1): 117-121. GAO C C, ZHAO C Y, WANG C, et al. Soil physicochemical properties and infiltration characteristics under different vegetation types in Tianlaochi watershed in the upstream of Heihe River[J]. Journal of Soil and Water Conservation, 2016, 30(1): 117-121. [31] LADO M, PAZ A, BEN-HUR M. Organic matter and aggregate-size interactions in saturated hydraulic conductivity[J]. Soil Science Society of America Journal, 2004, 68(1): 234-242. [32] XIAO B, MA S, HU K L. Moss biocrusts regulate surface soil thermal properties and generate buffering effects on soil temperature dynamics in dryland ecosystem[J]. Geoderma, 2019, 351: 9-24. [33] CASTER J, SANKEY T T, SANKEY J B, et al. Biocrust and the soil surface: Influence of climate, disturbance, and biocrust recovery on soil surface roughness[J]. Geoderma, 2021, 403: e115369. [34] HU W, SHAO M G, WANG Q J, et al. Temporal changes of soil hydraulic properties under different land uses[J]. Geoderma, 2009, 149(3/4): 355-366. [35] ZHU P Z, ZHANG G H, YANG Y, et al. Infiltration properties affected by slope position on cropped hillslopes[J]. Geoderma, 2023, 432: e116379. [36] 王浩, 张光辉, 刘法, 等. 黄土丘陵区生物结皮对土壤入渗的影响[J]. 水土保持学报, 2015, 29(5): 117-123. WANG H, ZHANG G H, LIU F, et al. lmpact of biological crust on soil infiltration in hilly areas of Loess Plateau[J]. Journal of Soil and Water Conservation, 2015, 29(5): 117-123. [37] BELNAP J, WILCOX B P, VAN SCOYOC M W, et al. Successional stage of biological soil crusts: An accurate indicator of ecohydrological condition[J]. Ecohydrology, 2013, 6(3): 474-482.
